

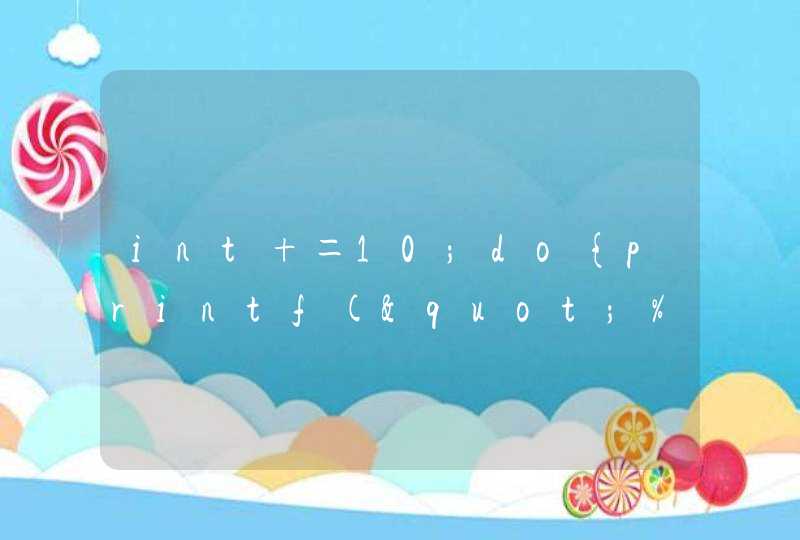






































有人这么描述转座子:基因组从出现就一直在经历一场和转座子及逆转录病毒的战争。真核生物的基因组到处是这场战争留下的疤痕,比如人类的基因组有45%的内容都跟转座子有关。
起源问题留给更专业的人来回答,要说到存在的意义,我来讲几个植物里的故事吧。
1. 转座子和孟德尔的豌豆
大家应该都记得孟德尔豌豆的黄圆绿皱吧?其中「皱」这个性状就和转座子有关,这个研究在1990年1月发表于《Cell》杂志「2」。我们当年再学高中生物的时候,一般把这个位点称为R/r, 它代表「皱」(rugosus), 它是隐形的,所以只有当两个等位基因都是r的时候,表型才会是「皱」。控制这个性状的基因是一个淀粉分支酶 (starch-branching enzyme, SBEI),这个基因被克隆后,发现在表型为「皱」的rr植物里,这个基因都有一个0.8kb的转座子插入,这个转座子和玉米里的Ac/Ds比较类似。这个插入就会导致淀粉分支酶SBEI失活,从而影响到种子里淀粉,脂类,以及蛋白质的代谢, 最终导致了「皱」的表型。
黑色部分就是转座子的插入。
2. 转座子与酿酒葡萄:卡本内 (Cabernet),霞多丽(Chardonnay)和奥山红宝石(Ruby Okuyama)
葡萄的颜色主要是由花青素 (anthocyanins)决定的,而花青素的合成主要由一些Myb相关基因控制,比如VlmybA1-1, VlmybA1-2, VlmybA2。关于葡萄颜色的研究又是一篇Science
我们把紫色的卡本内 (Cabernet)当作野生型来看的话,一个叫做Gret1的LTR反转录转座子插入了这个Vvmby1A基因,这个功能缺失突变就导致了绿葡萄的产生,比如霞多丽 (Chardonnay)。 之后的一次Gret1的重排事件导致了表型的部分恢复,这个事件就导致了另一种红颜色葡萄的出现,比如奥山红宝石 (Ruby Okuyama)。
制备目的基因主要依靠基因合成和基因分离两种方法;
目的基因获得方法是取得含有所需生物学功能或编码所需产物结构基因的DNA片段的方法。1969年贝克威思(J.Beckwith)等从大肠杆菌中分离出了乳糖操纵子DNA,以后有不少科学家用生物学方法、物理化学方法分离出许多纯化的基因。但总的来看为数不多。
1970年首次完成了77对碱基的酵母丙氨酸tRNA基因的全合成1973年又合成了126对碱基的酪氨酸tRNA的结构基因。但上述两次合成的基因都不能得到表达。1976年8月成功地合成了能表达功能的人工基因——193对碱基的大肠杆菌酪氨酸tRNA基因。自此以后人工合成基因的报道不断出现。
扩展资料:
目的基因相关的操作包括目的基因鉴定、分离、克隆、转化及转化后鉴定和测试等。通常,目的基因要么是已知供体生物基因组“文库”的一部分,要么是来自于先前未研究过的供体生物基因组。无论是哪种情况,聚合酶链式反应(PCR)技术都可以是目的基因倍增至生成构建体所需的数百万拷贝的水平。
当产生许多拷贝的目的基因后,可以通过酶切反应(或gataway系统),将目的基因酶促连接到构建载体中,然后将整个构建在细菌中表达,从报告基因选取阳性转化体,通常涉及含有插入DNA的菌落中的一些颜色变化。挑取阳性菌落,做液体培养,提取质粒DNA,进行酶切,切胶纯合。获得的DNA可用于后续的转化。




























